My first post on this blog was about fixation of a new beneficial mutation. I presented Haldane’s original 1927 result showing that with some reasonable assumptions, the likelihood that a new mutation with a small selective advantage 

The specific probability distribution for the number of “children” (i.e., new infected hosts) of a particular organism is assumed to be Poisson by Haldane. If you recall, we were evaluating the fixed point of the generating function of the distribution. The generating function of the Poisson distribution is
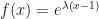
We then solved the equation 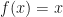


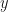
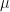
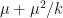
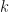
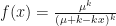
The goal is then to find a solution to the fixed point

This can be simplified to give an approximate solution valid for an arbitrary distribution:
![x \approx exp \left [ \frac{2(1 - \mu)}{\sigma^2} \right ]](https://s0.wp.com/latex.php?latex=x+%5Capprox+exp+%5Cleft+%5B+%5Cfrac%7B2%281+-+%5Cmu%29%7D%7B%5Csigma%5E2%7D+%5Cright+%5D&bg=ffffff&fg=111111&s=0&c=20201002)
Applied to the Poisson distribution where the mean and sigma are both 
![y = 1 - x \approx 1 - exp \left [\frac{-2s}{1 + s} \right ] = 2s - 4s^2 + \frac{22s^3}{3} \ldots](https://s0.wp.com/latex.php?latex=y+%3D+1+-+x+%5Capprox+1+-+exp+%5Cleft+%5B%5Cfrac%7B-2s%7D%7B1+%2B+s%7D+%5Cright+%5D+%3D+2s+-+4s%5E2+%2B+%5Cfrac%7B22s%5E3%7D%7B3%7D+%5Cldots&bg=ffffff&fg=111111&s=0&c=20201002)
And we see that we recapitulate Haldane’s result with the first term of the Taylor series approximation.
Using the negative binomial distribution, we get
![y = 1 - x \approx 1 - exp \left [\frac{-2s}{1 + 1/k + s + 2s/k + s^2/k}\right ]](https://s0.wp.com/latex.php?latex=y+%3D+1+-+x+%5Capprox+1+-+exp+%5Cleft+%5B%5Cfrac%7B-2s%7D%7B1+%2B+1%2Fk+%2B+s+%2B+2s%2Fk+%2B+s%5E2%2Fk%7D%5Cright+%5D&bg=ffffff&fg=111111&s=0&c=20201002)
Assume
![y \approx 1 - exp \left [\frac{-2s}{1 + 1/k + s + 2s/k}\right] = \frac{2s}{1 + 1/k} - \frac{4s^2}{1 + 1/k} \ldots](https://s0.wp.com/latex.php?latex=y+%5Capprox+1+-+exp+%5Cleft+%5B%5Cfrac%7B-2s%7D%7B1+%2B+1%2Fk+%2B+s+%2B+2s%2Fk%7D%5Cright%5D+%3D+%5Cfrac%7B2s%7D%7B1+%2B+1%2Fk%7D+-+%5Cfrac%7B4s%5E2%7D%7B1+%2B+1%2Fk%7D+%5Cldots&bg=ffffff&fg=111111&s=0&c=20201002)
And we see that the introduction of overdispersion by the negative binomial decreases the likelihood of fixation of the variant by approximately a factor of 
